

レプリカ交換分子動力学法による
アミロイド前駆体蛋白質の膜貫通部位の二量体構造予測


分子スケール研究開発チーム
宮下 尚之(左)
理化学研究所 基幹研究所(分子スケールWG)
杉田 有治(右)
アルツハイマー病は神経変性疾患の一つで認知機能の低下を主な症状とします。この病気の進行過程では脳神経細胞(ニューロン)死が生じますが、アミロイド前駆体蛋白質(APP)の断片であるアミロイドベータ(Aβ)ペプチドが凝集化し脳に蓄積されることが引き金になるというアミロイド仮説が最も有力な説として知られています。従って、Aβペプチドの生成に関する知見を深める事はアルツハイマー病を理解する上できわめて重要です。
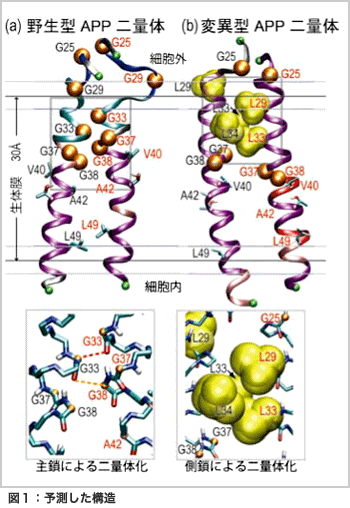
APPは約700個(残基)のアミノ酸でできておりニューロン表面の生体膜に存在します。Aβペプチドは、APPの細胞外から膜中に至る約40残基の部分で、上下2カ所がβ切断酵素とγ切断酵素という酵素によって順に切断されることにより生成されます。Aβペプチドとしては、40残基のアミノ酸でできているAβ1-40が最も多く生成されますが、γ切断部位に幅がある為にしばしば老人班の主要成分であるAβ1-42を生じます。私たちは異なる長さのAβペプチドがどのような分子機構で生体膜中から生じるのかを理解したいと考えました。しかし、APPの膜貫通部分の構造は生体膜中で大きく揺らいでいるせいか、未だにX線結晶構造解析によって決定された構造はありません。そこで、計算機を用いた分子シミュレーションを行うことによって、その構造を予測し、生化学実験データなどと比較することにしました。
本研究でターゲットとしたのは、APPの生体膜付近の重要な部分(Aβ23-55)です。この部分には5つのグリシン(Gly)が並んでおり、生体膜中でペア(二量体)を作る可能性が示唆されていました。実際、野生型(通常のアミノ酸配列を持つ)のAPPは生体膜中で二量体を形成していることが近年実験的に明らかにされました。ところが、興味深いことに5つのGlyのうちで2つのGly(Gly29とGly33)を疎水性残基(注)であるロイシン(Leu)に置き換えると(変異型)、野生型と同様に二量体を形成するにも関わらず、γ切断酵素によって切断されなくなる(すなわち、Aβ ペプチドが生成されなくなる)ことが報告されました[1]。たった2残基のアミノ酸変異がAPPの構造や機能にどのような影響を与えたのでしょうか?
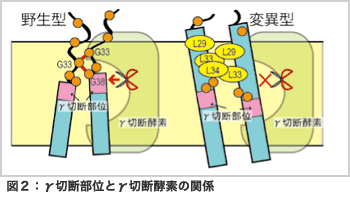
このような問いに答えるためには、通常、蛋白質内の分子間相互作用を考慮した分子動力学計算が有効です。しかし、X線結晶構造解析によって実験的に立体構造が得られていない蛋白質に関しては、多くの立体構造を計算機上で探索して得られた熱力学的に安定な構造を元に議論する必要があります。本研究で用いたレプリカ交換分子動力学法[2]をはじめとする拡張アンサンブル法はそのような目的にきわめて適した方法であり、これまでに名古屋大学の岡本祐幸教授をはじめとする日本国内の研究者が多くの新しい方法を開発してきました。我々の計算結果[3]は、図1に示すようにAPPの野生型と変位型では異なる二量体を形成すること、変位型では疎水性アミノ酸残基の数が増えたことによりAPPが膜に対してより傾いた構造になりAPPの切断部位がγ切断酵素の活性部位とずれてしまうことを示唆しました[図2]。この計算結果は、これまでに得られている実験データをよく再現するだけでなく、新しい分子機構を提案したことにより国内外の多くの実験家からも興味を持たれています。
本研究では、現在利用可能な計算機能力の限界により、生体膜の寄与を露わに取り込むことをせず排除体積効果を用いて水や膜の効果を陰的に取り扱ったモデルを用いました。しかし、次世代スーパーコンピュータが利用可能になると、生体膜を構成する脂質分子を露わに取り込んだ計算が可能になり、より信頼性の高い結果が得られると期待されます。また、本研究で用いたレプリカ交換法を始めとする拡張アンサンブル法は、利用可能なCPUの数が増えれば増えるほど効率良い計算が可能になってくるため、より大きな蛋白質の構造変化や基質結合に関する計算が実現できるはずです。アルツハイマー病のみならず多くの疾患に関わる現象が生体膜中に存在する蛋白質の挙動と関わることが知られていますので、我々の研究が将来、様々な疾患に関わる生命現象の理解に繋がり、いずれ病気の治療にも役立つことを期待しています。
(注)蛋白質を構成する20種類のアミノ酸は中心炭素原子(Cα)を持ち、その炭素原子に水素原子、アミノ基(NH2)、カルボキシル基(COOH)が結合した主鎖と呼ばれる共通構造を持っている。アミノ酸の違いはCα炭素原子に結合している側鎖で区別される。20種類の側鎖は水を好む親水性残基、水を嫌う疎水性残基に分類できる。
参考文献
[1] P. Kienen-Campard et al., J. Biol. Chem . 283, 7733-7744, (2008)
[2] Y. Sugita and Y. Okamoto, Chem. Phys. Lett ., 314, 141-151, (1999)
[3] N. Miyashita, J. E. Straub, D. Thirumalai and Y. Sugita, J. Am. Chem.
Soc ., 131, 3438-3439, (2009)
BioSupercomputing Newsletter Vol.1
- SPECIAL INTERVIEW
- 生命現象の本質に迫る革新的なアプローチ バイオスーパーコンピューティングによる挑戦が始まる
次世代計算科学研究開発プログラム 副プログラムディレクター 姫野 龍太郎
- LEADER’S TALK
- 生命活動の基礎となる生体高分子が担う機能をシミュレーションによってとらえる
分子スケール研究開発チーム チームリーダー 木寺 詔紀 - 三次元的に人体の全身モデルを構築して生体内で起こる現象を理解し、医療に役立てる
臓器全身スケール研究開発チーム チームリーダー 高木 周 - 第4の方法論「データ解析融合」によってバイオロジーを予測可能な科学へと導く
データ解析融合研究開発チーム チームリーダー 宮野
- 研究報告
- レプリカ交換分子動力学法によるアミロイド前駆体蛋白質の膜貫通部位の二量体構造予測
分子スケール研究開発チーム 宮下 尚之 /理化学研究所 基幹研究所 杉田 有治 - 重粒子線治療シミュレーション
臓器全身スケール研究開発チーム 石川 顕一 - ゲノムワイド関連解析と遺伝的、非遺伝的要因による治療予後予測の展望
理化学研究所ゲノム医科学研究センター 鎌谷 直之 - ペタスケールコンピューティングを支える基盤技術
生命体基盤ソフトウェア開発・高度化チーム 小野 謙二 / 伊東 聰 / 渡邉 大介
- ワークショップ報告
- VPHとのジョイントワークショップを開催