

Toward a spiking neuron-level model
of the early saccade visuomotor system

Kyoto University Jan Morén (photo)
Nara Institute of Science and Technology Tomohiro Shibata
Okinawa Institute of Science and Technology Kenji Doya
(Brain and Neural Systems WG)
Macroscopic visuomotor behaviour depends on high level interaction among system-scale structures in the brain. The functional mechanisms of such structures are however rooted in the low-level neurophysiology. To fully understand organic behaviour we need to look at these structures at both scales. Multiple description levels rapidly increase both the structural complexity and computational demands of the resulting model.
Our ultimate goal is to create the entire perception-action loop, from retina to the eye muscle motor systems, by ourselves and by collaboration with other groups, and we currently aim at neuron-level modeling for generating saccadic eye movements (saccades) with a particular emphasis on superior colliculus (SC), as saccades are a good target in that they are highly repeatable and stereotypical.
We use the NEST simulation tool to create a large-scale spiking-neuron level model of the intermediate SC (SGI)(Fig. 1), based on the present best knowledge on neurophysiology. We use the conductance-based Adaptive Exponential IaF neuron model augmented with an NMDA synaptic input. The number of neurons in the model is on the order of 100k, with burst, buildup, quasivisual, deep inhibitory and inhibitory interneurons, as well as supporting units for input generation and activity integration. The SGI is retinitopic and forms the first part of the oculomotor system. The retina projects to the cortical vision systems but also to the superficial SC. It in turn selects locally salient areas and activates the corresponding point in the intermediate division directly and through indirect pathways. Whereas the superficial division is a sensory area, the intermediate division is part of the motor system; they make up the sensor-motor convergence point in this perception-action loop.
A saccade is initiated in the SGI with buildup neuron activation spreading around the salient point. The lateral SGI is normally inhibited by neurons in the brainstem and by other areas, simplified here to a single inhibitory input. With the release of external inhibition, burst neurons generate an NMDA synapse-mediated burst of spikes that activates burst neuron systems in the pons and midbrain. They in turn move the eye horizontally and vertically, regulated by the cerebellum.
The SGI is fairly well understood but there are still a number of questions surrounding the nature of the activity in this area. The bursts may be a retinotopic target indicator, a movement vector or a velocity command. The significance of the spreading activation among buildup neurons is also not clear. We attempt to account for recent results on the spiking neuron activity and the spreading activation among buildup neurons, while keeping with the known neurophysiology of this area (Fig. 2).
Burst neuron activity indicates a saccade distance-dependent eye movement profile over time. The bursting profile is created through NMDA synapse-generated bursts with linear inhibitory feedback from a spike accumulator in the mesencephalic reticular formation. The resulting activity follows experimentally determined bursting profiles. Buildup neuron activity spread is accomplished by local intraconnections and inhibitory interneruons. We propose that the amount, but not the position, of the activity tracks saccade progess over time. This triggers inhibitory neurons in the deep SC that in turn inhibit burst neuron activity directly, and indirectly restores control to the rostral pole. This improves saccade end timing accuracy.
We aim to create a superficial SC and to pair our model to similarly detailed models of the retina and brainstem systems. We are also exploring embodiment by connecting the model to physical systems, enabling us to study computationally heavy model behavior in more naturalistic settings.
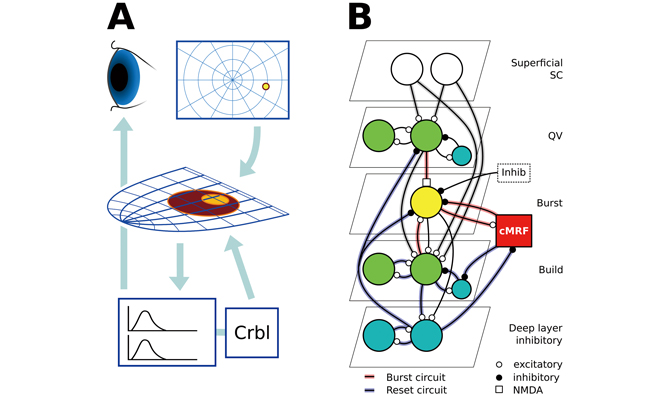
Fig. 1 : A) The main components and flow of the subcortical saccadic system.Retinal output activates the retinitopic SC, which in turn activates horizontal andvertical eye movement systems in the pons and the midbrain, and the cerebellumacting as a regulator.
B) The intermediate SC (SGI) model circuit. Red connections form the burst generation circuit, and blue connections the spreading activation and saccade system reset. Grey connections are common to both.
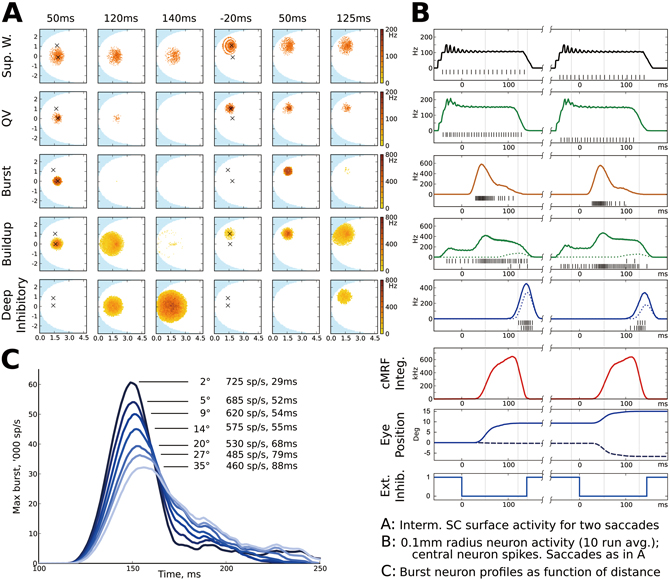
Fig. 2 : A) SGI surface activity over time for a 9 ° radius, 0 ° angle, then a 8 ° radius, 45° angle saccade. Each point is a 10ms activity average of one model neuron. Times relative to saccade deinhibition. From the top, superficial wide-field neurons; quasivisual neurons; burst neurons; buildup neurons and deep inhibitory neurons. From left, burst peak, buildup activity peak, deep inhibitory reset; preparatory activity for second saccade, second burst peak and second buildup peak.
B) 10-trial averaged activity in a 0.1mm radius around the central point for each saccade in A), in the same row order. black bars are spikes of the centermost neuron during a single trial, upper row for first saccade, lower row for second. Dotted lines for each saccade shows activity at the point of the other saccade. Below, total cMRF integrator activity; estimated horizontal (solid) and vertical (hashmarks) eye position; and external inhibition.
C) SGI burst neuron activity over time as a function of saccade distance. Longer saccades have a lower activity peak but a longer duration.
BioSupercomputing Newsletter Vol.6
- SPECIAL INTERVIEW
- Development of New Fluid-structure Interaction Analysis (ZZ-EFSI) Resulting in Rapid Achievement of High Operation Performance
Research Associate Professor, School of Engineering, The University of Tokyo Kazuyasu Sugiyama - Interview with High-performance Computing Team Members: Continued Efforts in Tuning to Harness the Potentials and the High Capability of the K Computer
Group Director of Computational Molecular Design Group,
Quantitative Biology Center, RIKEN
Makoto Taiji
Senior Researcher of High-performance Computing Team,
Integrated Simulation of Living Matter Group,
Computational Science Research Program, RIKEN
Yousuke Ohno
Senior Researcher of High Performance Computing Development Team,
High Performance Computing Development Group,
RIKEN HPCI Program for Computational Life Sciences
Hiroshi Koyama
Researcher of High-performance Computing Team,
Integrated Simulation of Living Matter Group,
Computational Science Research Program, RIKEN
Gen Masumoto
Research Associate of High-performance Computing Team,
Integrated Simulation of Living Matter Group,
Computational Science Research Program, RIKEN
Aki Hasegawa
- Report on Research
- Functional Analysis of Multidrug Efflux Transporter AcrB by All-Atom Molecular Dynamics Simulation
Graduate School of Nanobioscience,
Yokohama City University
Tsutomu Yamane,
Mitsunori Ikeguchi
(Molecular Scale WG) - Multi-scale Modeling of the Human Cardiovascular System
Computational Science Research Program,
RIKEN Liang Fuyou (Organ and Body Scale WG) - Toward a spiking neuron-level model of the early saccade visuomotor system
Kyoto University Jan Moren
Nara Institute of Science and Technology Tomohiro Shibata
Okinawa Institute of Science and Technology Kenji Doya
(Brain and Neural Systems WG) - Developing the MD Core Program for Large Scale Parallel Computing
Computational Science Research Program,
RIKEN Yousuke Ohno (High-performance Computing Team)
- SPECIAL INTERVIEW
- Pioneering the Future of Computational Life Science toward Understanding and Prediction of Complex Life Phenomena
Program Director of RIKEN HPCI Program for Computational Life Sciences
Toshio Yanagida
Deputy- Program Director of RIKEN HPCI Program for Computational Life Sciences
Akinori Kidera
Deputy- Program Director of RIKEN HPCI Program for Computational Life Sciences
Yukihiro Eguchi
- Report on Research
- Free Energy Profile Calculations for Changes in Nucleosome Positioning with All-Atom Model Simulations
Quantum Beam Science Directorate, Japan Atomic Energy Agency
Hidetoshi Kono, Hisashi Ishida, Yoshiteru Yonetani, Kimiyoshi Ikebe (Field 1- Program 1) - Estimation of Skeletal Muscle Activity and Neural Model of Spinal Cord Reflex
Information Science and Technology, The University of Tokyo
Yoshihiko Nakamura (Field1 - Program 3)
- Report
- ISLiM Interim Accomplishment Meeting in 2011
Computational Science Research Program, RIKEN Eietsu Tamura - Computational Life Sciences Classes Held in High Schools
HPCI Program for Computational Life Sciences, RIKEN
Chisa Kamada, Yasuhiro Fujihara, Yukihiro Eguchi