

アクチン細胞骨格の計算メカノバイオロジー

京都大学再生医科学研究所
井上 康博(細胞スケールWG)
アクチン細胞骨格は、およそ5nm程度の大きさのアクチン単量体が互いに繊維状に連結したアクチンフィラメントを基本構造に、様々な補助タンパク質との相互作用を介して、網目構造や束構造などの高次構造を作り、細胞の形態や運動を支える重要な力学的・化学的基盤を成しています。例えば、細胞表面のアクチンは、糸状仮足や葉状仮足などの運動性を担う構造を作り、これらは、複雑に枝分かれしたニューロン構造の形成や創傷治癒における細胞運動などに必要です。また、アクチンとミオシンからなる複合繊維(アクトミオシン)構造では、ミオシンのモーター運動により、力が発生し、形態形成における細胞変形や収縮環による細胞質分裂、細胞接着のターンオーバーなどにおいて重要となります。これら構造は、アクチンを中心とした分子システムの自己組織化によって現れ、また、常に力学的・化学的に準安定な状態を経巡ることで、ダイナミックにその構造や特性を変化させることができます。私たちは、このようなアクチン細胞骨格のダイナミクスと細胞機能との関連について、計算メカノバイオロジーの手法を用いて研究しています。最近の私たちの研究により、細胞運動における細胞形状とアクチン重合が熱ゆらぎを介して連成すること[1]や、アクトミオシンネットワークに発生する力とミオシン濃度との関係がネットワーク構造の変化に対応して双線形になること[2]を明らかにしました。これらは、細胞スケールの機能発現と分子スケールのダイナミクスとの関係について、重要な知見を与えています。また、計算機を用いてアクチンメカノバイオロジーシミュレーションができるようになってきたことで、細胞をまるごと1個、計算機上に再現することも全く手の届かない夢ではなくなってきました。もちろん、まだ、解決しなければならない課題は山積みです。これらに関しては、次世代生命体統合シミュレーション研究グループ内外の研究者と連携する必要があり、現在、そのような場として、計算科学アルゴリズム研究会が定期的に開催され、分野を超えて数理モデル・計算手法の知見を共有し、また、関連する境界領域の研究者との連携が進められています。
【参考文献】
[1] Inoue, Y. and Adachi, T. Biomech. Model. Mechanobiol. 2011 10:495-503.
[2] Inoue, Y. Tsuda, S., Nakagawa K., Hojo M., Adachi T. J Theor Biol. 2011 21;281(1):65-73.
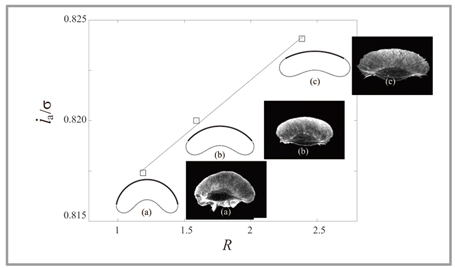 |
図1:移動性細胞の計算機シミュレーション:シミュレーションによって得られた細胞形状(実線図)は、実際に観察される細胞形状(蛍光画像)と一致しました。また、シミュレーションにより、細胞先端のアクチン重合速度(縦軸)は、細胞先端の曲率半径(横軸)に比例することがわかりました。 |
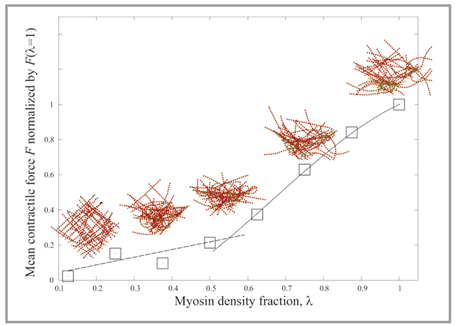 |
図2:アクトミオシンネットワークの自律的構造変化:アクトミオシンネットワークに存在するミオシンのモーター運動によって、アクトミオシンネットワークの構造は自律的に変化します。シミュレーションにより、構造再編に伴い、ネットワークに発生する力は、ミオシン分率に対して双線形となることがわかりました。 |
BioSupercomputing Newsletter Vol.5
- SPECIAL INTERVIEW
- “予測する生物学”をめざすバイオスーパーコンピューティングの挑戦はいよいよ世界一の「京」で成果を出すフェーズに入った
理化学研究所 次世代計算科学研究開発プログラム 副プログラムディレクター 姫野 龍太郎 - 高性能計算機資源および開発アプリケーションの産業利用促進を図るために何をすべきか
計算科学振興財団 チーフコーディネーター 福田 正大
都市活力研究所 主席研究員 バイオグリッドセンター関西 理事・事務局長 志水 隆一
- 研究報告
- QM/MM 自由エネルギー法による酵素反応分子機構の解析
京都大学大学院理学研究科 林 重彦(分子スケールWG) - アクチン細胞骨格の計算メカノバイオロジー
京都大学再生医科学研究所 井上 康博(細胞スケールWG) - 血栓シミュレーションに向けた血流解析手法の開発
東京大学工学系研究科 伊井 仁志(臓器全身スケールWG) - 生命体シミュレーションのためのデータ同化技術の開発
統計数理研究所 樋口 知之(データ解析融合WG)
- SPECIAL INTERVIEW
- 複雑な生命現象の理解と予測に向けて計算生命科学の明日を拓く
理化学研究所 HPCI計算生命科学推進プログラム プログラムディレクター 柳田 敏雄
理化学研究所 HPCI計算生命科学推進プログラム 副プログラムディレクター 木寺 詔紀
理化学研究所 HPCI計算生命科学推進プログラム 副プログラムディレクター 江口 至洋
- 研究報告
- 創薬応用シミュレーション
東京大学先端科学技術研究センター 藤谷 秀章(分野1-課題2) - 次世代DNAシークエンサデータの超高速解析
東京工業大学大学院情報理工学研究科
秋山 泰 / 石田貴士 / 角田将典 / 鈴木脩司(分野1-課題4)
- 報告
- バイオスーパーコンピューティングサマースクール2011
理化学研究所 次世代計算科学研究開発プログラム
石峯 康浩(臓器全身スケールWG)
統計数理研究所 データ同化研究開発センター
斎藤 正也(データ解析融合WG)
新潟国際情報大学
近山 英輔(細胞スケールWG)
東海大学医学部内科学系循環器内科
七澤 洋平(細胞スケール/臓器全身スケールWG)
理化学研究所 次世代計算科学研究開発プログラム
半田 高史(脳神経系WG)
理化学研究所 次世代計算科学研究開発プログラム
舛本 現(開発・高度化T)
理化学研究所 次世代計算科学研究開発プログラム
森次 圭(分子スケールWG) - 「次世代生命体統合シミュレーションソフトウェアの開発(ISLiM)」開発アプリケーション紹介ページ、オープン
次世代計算科学研究開発プログラム 次世代生命体統合シミュレーション研究推進グループ